 |
The
Muchhala Lab University of Missouri - St. Louis |
 |
| Home | People |
Research
|
Press | Bat-Flower Images | Publications | Links |
|
Does biodiversity beget biodiversity? Alfred Wallace suggested that high tropical diversity is due to the “complex influences of organism upon organism”: an increased number of biotic interactions increases the number of specialized niches available, accelerating adaptation and speciation in a positive feedback loop. The Muchhala Lab conducts research in evolutionary ecology addressing the role of such interactions, especially mutualism and interspecific competition, in structuring communities and driving diversification. We focus on plants pollinated by bats and hummingbirds, and integrate various approaches including molecular phylogenetics, mathematical modeling, and field experiments. Three specific questions we are particularly interested in include: 1) What selective pressures favor specialization in pollination systems? 2) What are the evolutionary and ecological consequences of competition for pollination? 3) How do pollinators influence plant speciation? |
|
| Specialization in Pollination Systems A common theme in angiosperm diversification involves repeated evolution
of suites of traits, termed pollination syndromes, that adapt a flower to a
certain type of pollinator ( Lagomarsino et al. 2017, Clark et al. 2015). But why
specialize to a given pollinator, when generalizing (e.g., Lagomarsino & Muchhala 2019) ought to allow many more
opportunities for successful pollination per unit time? Work with the genus Burmeistera
(Campanulaceae) showed that most species are specialized to either bats or
hummingbirds (Muchhala 2006a), and experiments demonstrated that an adaptive
tradeoff in floral form (wide flowers fit bat heads better while narrow flowers
fit hummingbird bills better) is strong enough that generalist flowers are
suboptimal (Muchhala 2007). We are
continuing to explore the traits that adapt flowers to bat pollination, such as
increased pollen production (e.g. Muchhala et al. 2010), and olfactory, visual,
and echo-reflectance cues (reviewed in Muchhala & Tschapka 2020). |
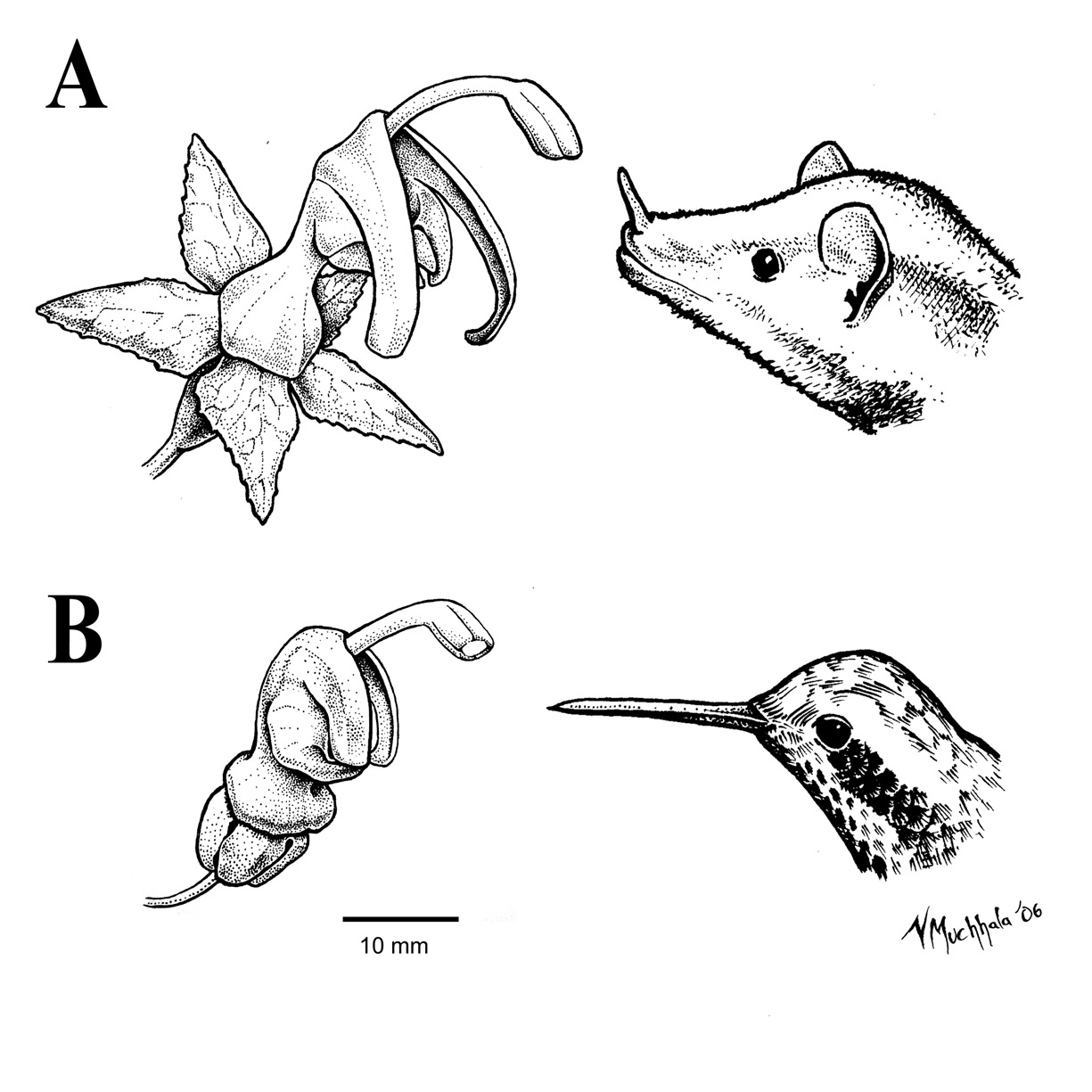 Burmeistera borjensis flower with Anoura geoffroyi (A) and B. rubrosepala with Adelomyia melanogenys (B) |
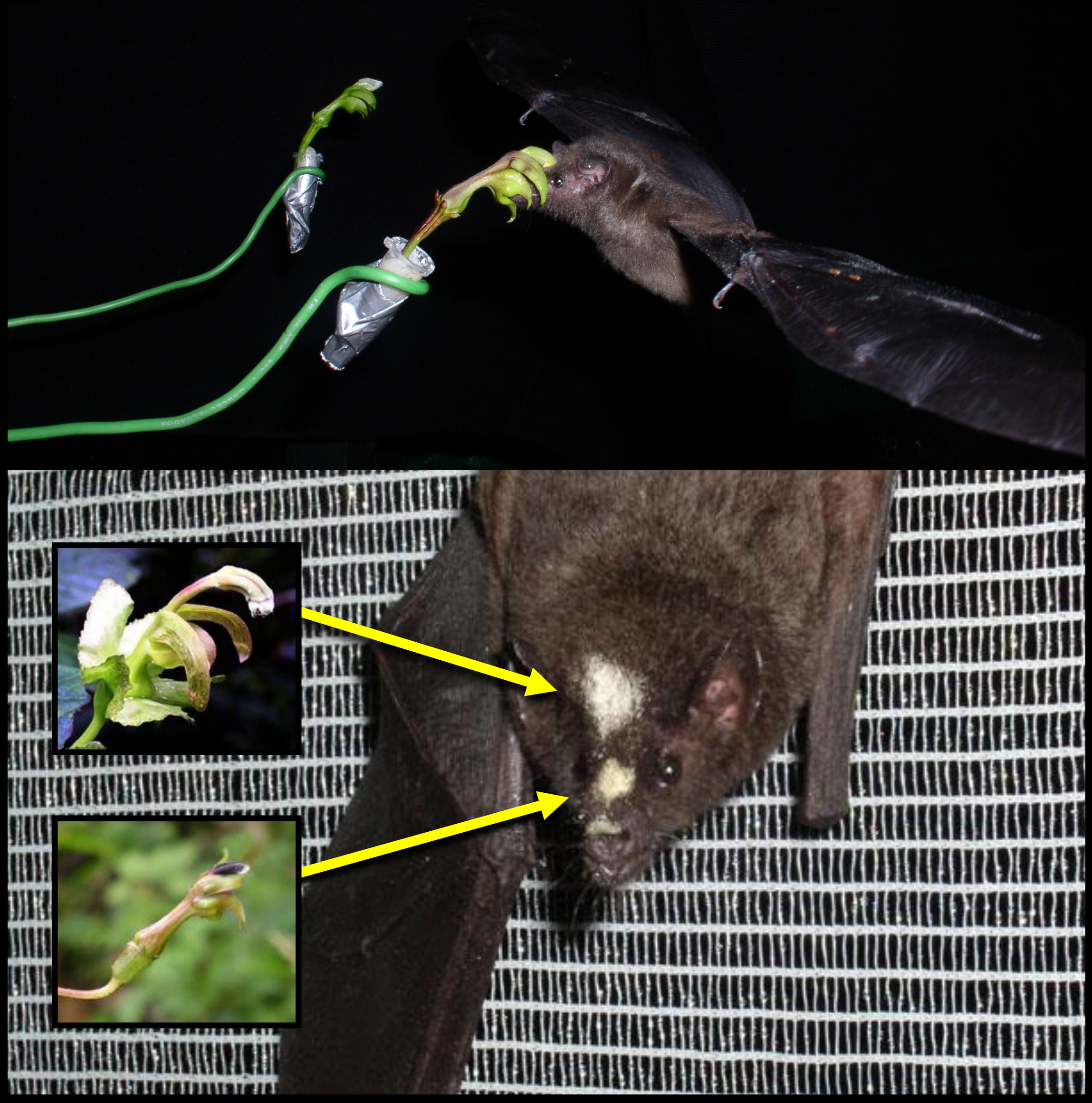
  Anoura fistulata drinking honey-water from a glass tube (top) and pollinating Centropogon nigricans (bottom) |
Specialization
in pollination systems (continued)
The majority of flowers are pollinated by more than one
species, and pollinators typically visit many species of flowers, thus
pollination
syndromes are examples of either one-sided evolution (plants evolving
to
pollinators) or, at best, diffuse coevolution. True reciprocal coevolution can only occur in
highly specialized interactions. One
example of this is the nectar bat Anoura
fistulata (Muchhala et al. 2005), which can launch its tongue 1.5 times its
body length (double that of other bats and longer than any other mammal).
Unique adaptations allow it to store its tongue in its rib cage (Muchhala
2006b). Experiments suggest that this
pair is involved in a coevolutionary race with the 9-cm-long flowers of
Centropogon nigricans; longer tongues allow bats to reach more nectar, while longer
flowers maximize pollen transfer (Muchhala & Thomson 2009). We plan to continue to study the geographic
mosaic of the interaction between this bat and the flowers it pollinates throughout
its range, as well as the ‘trigger’ behind the coevolutionary race – that is,
why exactly do long flowers receive more pollen? |
| Competition for pollination A given species of plant occurs within a community of many other plants. How does an individual ensure that it will receive conspecific pollen and that its pollen reaches conspecifics? We recently reviewed the literature to summarize our understanding of this topic (Moreira-Hernandez & Muchhala 2019). The costs of competition may include loss of pollen to foreign flowers (Muchhala & Thomson 2012) or blockage of stigmas with foreign pollen (Moreira-Hernandez et al. 2019). Hummingbird-pollinated Iochroma demonstrate one solution to this problem: co-occurring species evolve differences in flower color to encourage hummingbirds to stay faithful to one species rather than switching between species in their foraging bouts (Muchhala et al. 2014). Bat-pollinated Burmeistera demonstrate a another solution: co-occurring species have evolved different lengths of the staminal column, thus placing pollen on different regions of bat’s heads (Muchhala & Potts, 2007). Ongoing work in the lab is exploring whether pollen transfer between Burmeistera species results in hybridization, and thus introgression, and is further detailing the fitness costs for male and female reproductive functions. |
 Floral character displacement in pollen placement: pollen transfer experiments (top) and bat with pollen from by two different Burmeistera species (bottom) |
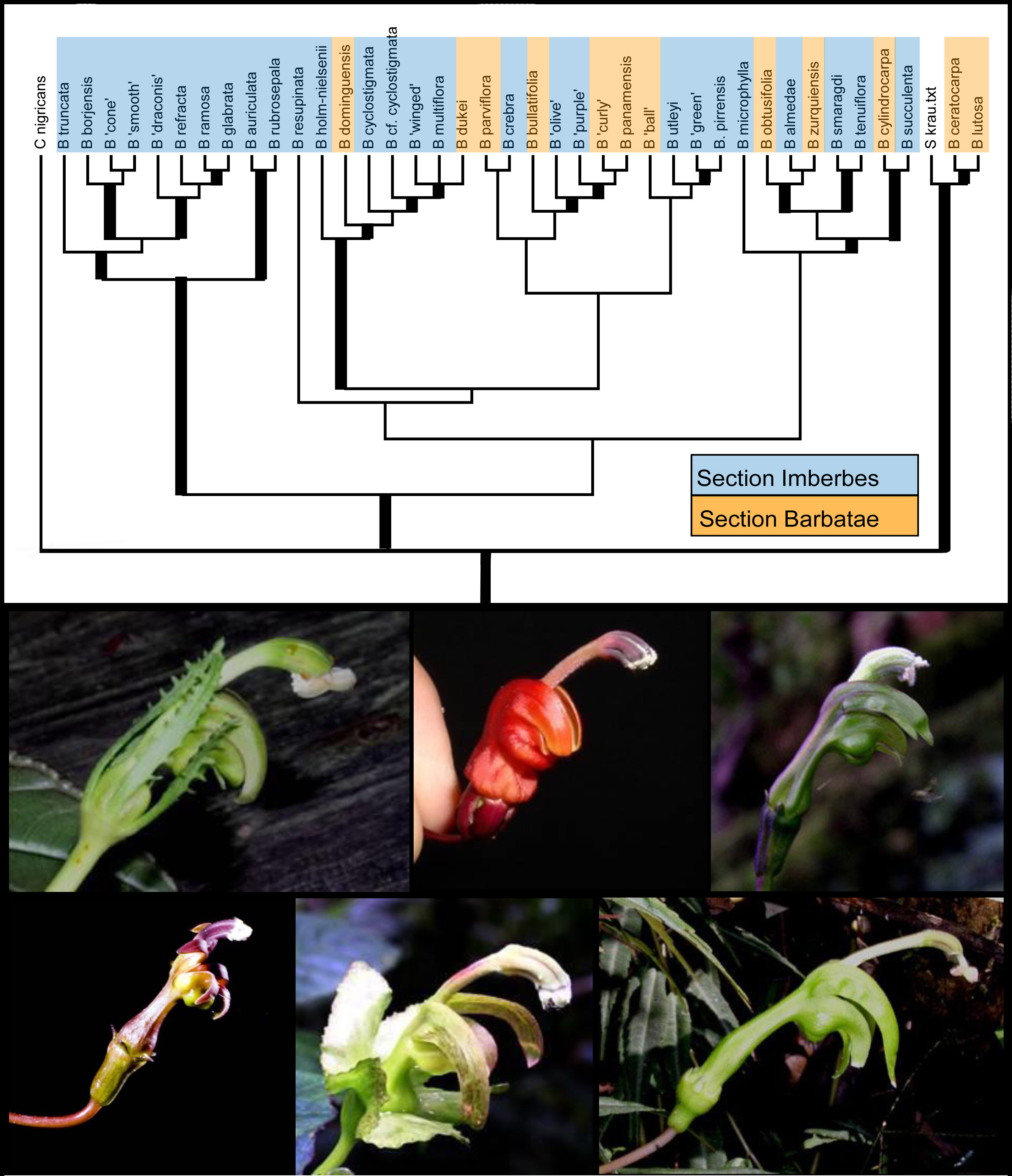 Preliminary phylogenetic hypothesis for Burmeistera (top) and a sampling of floral diversity across the genus (bottom) |
Plant speciation What evolutionary innovations allowed the remarkable and rapid radiation of flowering plants? One long-standing paradigm holds that coevolution with pollinators is the key (reviewed in Armbruster & Muchhala 2009), because speciation rates are directly amplified via pollinator-mediated reproductive isolation. Post-pollination processes have received much less attention. To begin to understand the relative importance of different isolating barriers for angiosperm speciation, they must be compared among multiple sister-species pairs of various ages; in this way we can distinguish those that directly contributed to speciation from those that only arose after speciation was complete. We are currently developing molecular phylogenies for Burmeistera (e.g. Uribe-Convers et al. 2017) and related genera (Lagomarsino et al. 2017), and will use these in combination with cross-pollination experiments to explore the importance of various pre- and post-zygotic reproductive barriers in the diversification of these clades. |
 Video of Anoura fistulata visiting Centropogon nigricans (© National Geographic Channel, 2012) |
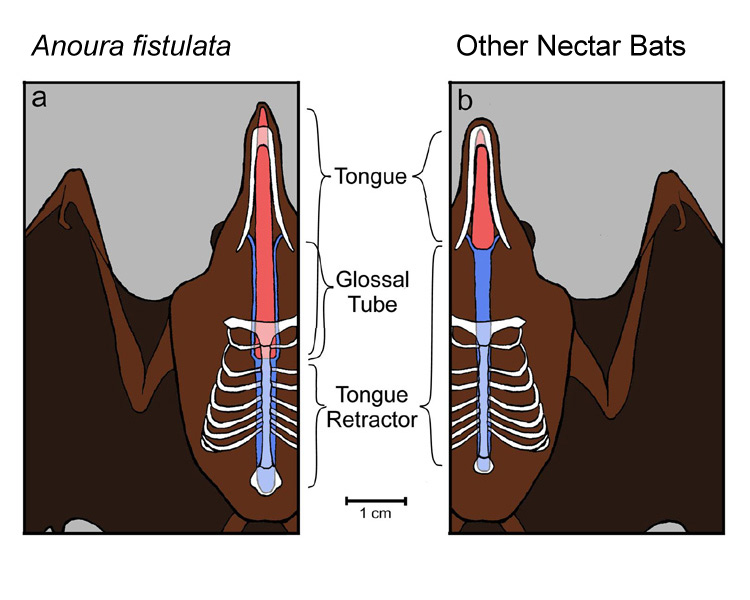 Comparison of the tongue morphology of A. fistulata and a typical glossophagine nectar bat |